Сердце костных рыб сколько камер в сердце
ГЛАВА I
СТРОЕНИЕ И НЕКОТОРЫЕ ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РЫБ
КРОВЕНОСНАЯ СИСТЕМА. ФУНКЦИИ И СВОЙСТВА КРОВИ
Главным отличием кровеносной системы рыб от других позвоночных является наличие одного круга кровообращения и двухкамерного сердца, наполненного венозной кровью (за исключением двоякодышащих и кистёперых).
Сердце состоит из одного желудочка и одного предсердия и помещается в околосердечной сумке, сразу за головой, позади последних жаберных дуг, т. е. по сравнению с другими позвоночными сдвинуто вперед. Перед предсердием имеется венозная пазуха, или венозный синус, со спадающими стенками; через эту пазуху кровь поступает в предсердие, а из него – в желудочек.
Расширенный начальный участок брюшной аорты у низших рыб (акулы, скаты, осетровые, двоякодышащие) образует сокращающийся артериальный конус, а у высших рыб – луковицу аорты, стенки которой сокращаться не могут. Обратному току крови препятствуют клапаны.
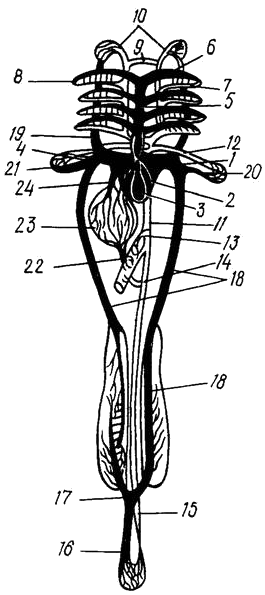
Схема кровообращения в самом общем виде представлена следующим образом. Венозная кровь, заполняющая сердце, при сокращениях сильного мускульного желудочка через артериальную луковицу по брюшной аорте направляется вперед и поднимается в жабры по приносящим жаберным артериям. У костистых рыб их четыре с каждой стороны головы – по числу жаберных дуг. В жаберных лепестках кровь проходит через капилляры и, окисленная, обогащенная кислородом, направляется по выносящим сосудам (их также четыре пары) в корни спинной аорты, которые затем сливаются в спинную аорту, идущую вдоль тела назад, под позвоночником. Соединение корней аорты спереди образует характерный для костистых рыб головной круг. Вперед от корней аорты ответвляются сонные артерии.
От спинной аорты идут артерии к внутренним органам и мускулатуре. В хвостовом отделе аорта переходит в хвостовую артерию. Во всех органах и тканях артерии распадаются на капилляры. Собирающие венозную кровь венозные капилляры впадают в вены, несущие кровь к сердцу. Хвостовая вена, начинающаяся в хвостовом отделе, войдя в полость тела, разделяется на воротные вены почек. В почках разветвления воротных вен образуют воротную систему, а выйдя из них, сливаются в парные задние кардинальные вены. В результате слияния вен задних кардинальных с передними кардинальными (яремными), собирающими кровь из головы, и подключичными, приносящими кровь из грудных плавников, образуется два Кювьерова протока, по которым кровь попадает в венозный синус. Кровь из пищеварительного тракта (желудка, кишечника) и селезенки, идущая по нескольким венам, собирается в воротную вену печени, разветвления которой в печени образуют воротную систему. Собирающая кровь из печени печеночная вена впадает прямо в венозный синус (рис. 21). В спинной аорте радужной форели обнаружена эластичная связка, выполняющая роль нагнетающего насоса, который автоматически увеличивает циркуляцию крови во время плавания, особенно в мускулатуре тела. Производительность этого ‛дополнительного сердца“ зависит от частоты движений хвостового плавника.
У двоякодышащих рыб появляется неполная перегородка предсердия. Это сопровождается и возникновением ‛лёгочного“ круга кровообращения, проходящего через плавательный пузырь, превращенный в легкое.
Сердце рыб относительно очень мало и слабо, гораздо меньше и слабее, чем у наземных позвоночных. Масса его обычно не превышает 0,33–2,5%, в среднем 1 % массы тела, тогда как у млекопитающих оно достигает 4,6%, а у птиц даже 10–16%.
Кровяное давление (Па) у рыб низкое – 2133,1 (скат), 11198,8 (щука), 15998,4 (лосось), тогда как в сонной артерии лошади – 20664,6.
Невелика и частота сокращений сердца – 18–30 ударов в минуту, причем она сильно зависит от температуры: при низких температурах у рыб, зимующих на ямах, она уменьшается до 1–2 ;у рыб, переносящих вмерзание в лед, пульсация сердца на этот период прекращается.
Количество крови у рыб относительно меньше, чем у всех остальных позвоночных животных (1,1 – 7,3% от массы тела, в том числе у карпа 2,0–4,7%, сома – до 5, щуки – 2, кеты – 1,6, тогда как у млекопитающих – 6,8% в среднем).
Это связано с горизонтальным положением тела (нет необходимости проталкивать кровь вверх) и меньшими энергетическими тратами в связи с жизнью в водной среде. Вода является гипогравитационной средой, т. е. сила земного притяжения здесь почти не сказывается.
Морфологическая и биохимическая характеристика крови различна у разных видов в связи с систематическим положением, особенностями среды обитания и образа жизни. Внутри одного вида эти показатели колеблются в зависимости от сезона года, условий содержания, возраста, пола, состояния особей.
Количество эритроцитов в крови рыб меньше, чем у высших позвоночных, а лейкоцитов, как правило, больше. Это связано, с одной стороны, с пониженным обменом рыб, а с другой – с необходимостью усилить защитные функции крови, так как окружающая среда изобилует болезнетворными организмами. По средним данным, в 1 мм3крови количество эритроцитов составляет (млн. ): у приматов –9,27; копытных– 11,36; китообразных – 5,43; птиц – 1,61–3,02; костистых рыб– 1,71 (пресноводные), 2,26 (морские), 1,49 (проходные).
Количество эритроцитов у рыб колеблется в широких пределах, прежде всего в зависимости от подвижности рыб: у карпа – 0,84–1,89 млн. /мм3 крови, щуки – 2,08, пеламиды – 4,12 млн. /мм3. Количество лейкоцитов составляет у карпа 20–80, у ерша – 178 тыс. /мм3. Клетки крови рыб отличаются большим разнообразием, чем у какой-либо другой группы позвоночных. У большинства видов рыб в крови имеются и зернистые (нейтрофилы, эозинофилы) и незернистые (лимфоциты, моноциты) формы лейкоцитов.
Среди лейкоцитов преобладают лимфоциты, на долю которых приходится 80–95%, моноциты составляют 0,5–11%; среди зернистых форм преобладают нейтрофилы–13–31%; эозинофилы встречаются редко (у карповых, амурских растительноядных, некоторых окуневых).
Соотношение разных форм лейкоцитов в крови карпа зависит от возраста и условий выращивания.
Общее количество лейкоцитов в крови рыб сильно изменяется в течение года, у карпа оно повышается летом и понижается зимой при голодании в связи со снижением интенсивности обмена.
Кровь окрашена гемоглобином в красный цвет, но есть рыбы и с бесцветной кровью. Так, у представителей семейства Chaenichthyidae (из подотряда нототениевых), обитающих в антарктических моряхв условиях низкой температуры (
Назад Оглавление
Оглавление Далее
Далее
КЛАСС РЫБЫ. Кровеносная система. Выделительная система. Хозяйственное значение рыб и охрана рыбных богатств.
Есть ли у рыбы сердце?
В кровеносной системе рыб, по-сравнению с ланцетниками, появляется настоящее сердце. Оно состоит из двух камер, т. е. сердце рыб двухкамерное.
Первая камера — это предсердие, вторая камера — это желудочек сердца. Кровь сначала попадает в предсердие, затем мышечным сокращением проталкивается в желудочек. Далее в результате его сокращения изливается в крупный кровеносный сосуд.
Сердце рыб находится в околосердечной сумке, расположенной за последней парой жаберных дуг в полости тела.
Как и у всех хордовы, кровеносная система рыб замкнутая.
Это значит, что нигде по пути своего следования кровь не покидает сосудов и не изливается в полости тела. Чтобы обеспечить обмен веществ между кровью и клетками всего организма, крупные артерии (сосуды, несущие кровь, насыщенную кислородом) постепенно ветвятся на более мелкие. Самые мелкие сосуды — капилляры. Отдав кислород и забрав углекислый газ, капилляры снова объединяются в более крупные сосуды (но уже венозные).
У рыб только один круг кровообращения.
При двухкамерном сердце по-другому быть и не может. У более высокоорганизованных позвоночных (начиная с земноводных) появляется второй (легочный) круг кровообращения. Но у этих животных и сердце трехкамерное или даже четырехкамерное.
Через сердце протекает венозная кровь, отдавшая кислород клеткам тела.
Далее эту кровь сердце толкает в брюшную аорту, который идет к жабрам и ветвится на приносящие жаберные артерии (но несмотря на название «артерии» они содержат венозную кровь). В жабрах (а конкретно, в жаберных лепестках) из крови в воду выделяется углекислый газ, а из воды в кровь просачивается кислород.
Происходит это в результате разницы в их концентрации (растворенные газы идут туда, где их меньше). Обогатившись кислородом, кровь становится артериальной. Выносящие жаберные артерии (уже с артериальной кровью) впадают в один крупный сосуд — спинную аорту.
Она проходит под позвоночником вдоль тела рыбы и от нее берут начало более мелкие сосуды. От спинной аорты также отходят сонные артерии, идущие к голове и снабжающие кровью в том числе головной мозг.
Перед тем как попасть в сердце венозная кровь проходит через печень, где очищается от вредных веществ.
В кровеносной системе костных и хрящевых рыб есть небольшие различия. В основном это касается сердца. У хрящевых рыб (и некоторых костных) расширенный участок брюшной аорты сокращается наряду с сердцем, а у большинства костных рыб — нет.
Кровь рыб красная, в ней присутствуют эритроциты с гемоглобином, связывающим кислород.
Однако эритроциты рыб имеют овальную форму, а не дисковидную (как, например, у человека). Количество крови, текущей по кровеносной системе, у рыб меньше, чем у наземных позвоночных.
Сердце рыб бьется не часто (около 20-30 ударов в минуту), и количество сокращений зависит от температуры окружающей среды (чем теплее, тем чаще).
Поэтому их кровь течет не так быстро и, следовательно, обмен веществ относительно медленный. Это, например, влияет на то, что рыбы — холоднокровные животные.
У рыб органами кроветворения являются селезенка и соединительная ткань почек.
Несмотря на то, что описанная кровеносная система рыб характерна для подавляющего большинства из них, у двоякодышащих и кистеперых она несколько отличается.
У двоякодышащих в сердце появляется неполная перегородка и появляется подобие легочного (второго) круга кровообращения. Но этот круг проходит не через жабры, а через плавательный пузырь, превращенный в легкое.
Через сердце рыб проходит а) артериальная кровь б) смешанная кровь в) венозная кровь?
Как выглядит сердце рыбы

Сердце рыбы щуки фото. Есть ли у рыбы сердце, конечно есть.
Фото рыбы щуки с сердцем. Кровь в сердце рыб проходит также как и у других обеспечивая органы всем для жизни не обходимым. Сколько сердец у рыбы, речной только одно.
Где у рыбы сердце, в районе гортани и у щуки оно продолжает биться не которое время даже после того как будет извлечено из рыбы. Какая кровь в сердце рыб, кровь в сердце рыбы щуки такого же красного цвета которая заметно темнеет при чистке.
Фото кровь в сердце рыбы. Полезные рыбы для сердца практически все речные, только размеры самого сердца слишком малы для употребления в гастрономических целях.
Эта запись была размещена в рыбы автор Vladimir Veselov (постоянная ссылка).
Кровь выполняет многочисленные функции только тогда, когда движется по сосудам. Обмен веществ между кровью и другими тканями организма происходит в капиллярной сети. Отличаясь большой протяженностью и разветвленностью, она оказывает большое сопротивление току крови. Давление, необходимое для преодоления сопротивления сосудов, создается в основном сердцем, Строение сердца рыб проще, чем высших позвоночных. Производительность сердца у рыб как нагнетательного насоса значительно ниже, чем у наземных животных.
Тем не менее оно справляется со своими задачами. Водная среда создает благоприятные условия для работы сердца. Если у наземных животных значительная часть работы сердца затрачивается на преодоление сил гравитации, вертикальные перемещения крови, то у рыб плотная водная среда существенно нивелирует гравитационные влияния.
Вытянутое в горизонтальном направлении тело, небольшой объем крови, наличие только одного крута кровообращения дополнительно облегчают функции сердца у рыб.
Классификация
В зоологической литературе XX века группа костных рыб обычно рассматривалась как класс
, состоящий из двух подклассов: Лучепёрые (Actinopterygii) и Лопастепёрые (Sarcopterygii)[10]. Такая трактовка сохраняется и в некоторых учебниках начала XXI века[11], однако всё большее распространение получают классификации, в которых ранг упомянутых подклассов повышен до ранга класса. Например, в классификации канадского зоолога Дж. С. Нельсона (2006) группа костных рыб (выступающая под названием Euteleostomi и не имеющая ранга) трактуется как объединение двух сестринских групп — классов Лучепёрые и Лопастепёрые. При этом если лучепёрые являются монофилетической группой, то лопастепёрые — группа заведомо парафилетическая (поскольку ближайший общий предок последней группы также является и предком современных четвероногих); в результате парафилетической оказывается и вся группа костных рыб. Следуя правилам кладистики, не допускающим использования парафилетических таксонов, Нельсон переопределяет объём лопастепёрых, включая в него и всех четвероногих (получивших в его классификации ранг инфракласса)[12].
Строение сердца рыб
Сердце у рыб небольшое, составляющее примерно 0,1% массы тела. Из этого правила, конечно, есть исключения. Например, у летучих рыб масса сердца достигает 2,5 % массы тела.
Для всех рыб характерно двухкамерное сердце. Вместе с тем существуют видовые различия в строении этого органа.
В обобщенном виде можно представить две схемы строения сердца в классе рыб. И в первом, и во втором случае выделяют 4 полости: венозный синус, предсердие, желудочек и образование, отдаленно напоминающее дугу аорты у теплокровных, — артериальную луковицу у костистых и артериальный конус у пластинчатожаберных (рис, 7.1). Принципиальное различие этих схем заключено в морфофункциональных особенностях желудочков и артериальных образований.
У костистых артериальная луковица представлена фиброзной тканью с губчатым строением внутреннего слоя, но без клапанов.
У пластинчатожаберных артериальный конус помимо фиброзной ткани содержит и типичную сердечную мышечную ткань, поэтому обладает сократимостью.
Конус имеет систему клапанов, облегчающих одностороннее продвижение крови через сердце.
Рис. 7.1. Схема строения сердца рыб
В желудочке сердца рыб обнаружены различия в структуре миокарда.
Принято считать, что миокард рыб специфичен и представлен однородной сердечной тканью, равномерно пронизанной трабекулами и капиллярами. Диаметр мышечных волокон у рыб меньше, чем у теплокровных, и составляет 6-7 мкм, что вдвое меньше по сравнению, например, с миокардом собаки. Такой миокард называют губчатым.
Сообщения о васкуляризации миокарда рыб довольно запутанны. Миокард снабжается венозной кровью из трабекулярных полостей, которые, в свою очередь, заполняются кровью из желудочка через сосуды Тибезия (Thebesian vessels). В классическом понимании у рыб нет коронарного кровообращения. По крайней мере, медики-кардиологи придерживаются такой точки зрения. Однако в литературе по ихтиологии термин «коронарное кровообращение рыб» встречается часто.
В последние годы исследователи обнаружили много вариаций васкуляризации миокарда. Например, С. Agnisola et. al (1994) сообщает о наличии двуслойного миокарда у форели и электрического ската. Со стороны эндокарда лежит губчатый слой, а над ним слой миокардиальных волокон с компактным упорядоченным расположением.
Исследования показали, что губчатый слой миокарда обеспечивается венозной кровью из трабекулярных лакун, а компактный слой получает артериальную кровь по гипобронхиальным артериям второй пары жаберных дут.
У elasmobranchs коронарное кровообращение отличается тем, что артериальная кровь из гипобронхиальных артерий доходит до губчатого слоя по хорошо развитой системе капилляров и попадает в полость желудочка по сосудам Тибезия.
Еще одно существенное различие костистых и пластинчатожаберных заключается в морфологии перикарда.
У костистых перикард напоминает таковой наземных животных. Он представлен тонкой оболочкой.
У пластинчатожаберных перикард образован хрящевой тканью поэтому он представляет собой как бы жесткую, но упругую капсулу.
В последнем случае в период диастолы в перикардиальном пространстве создается некоторое разрежение, что облегчает кровенаполнение венозного синуса и предсердия без дополнительных затрат энергии.
Электрические свойства сердца рыб
Строение миоцитов сердечной мышцы рыб сходно с таковым высших позвоночных.
Поэтому и электрические свойства сердца похожи. Потенциал покоя миоцитов у костистых и пластинчатожаберных составляет 70 мВ, у миксин — 50 мВ. На пике потенциала действия регистрируется изменение знака и величины потенциала с минус 50 мВ до плюс 15 мВ. Деполяризация мембраны миоцита приводит к возбуждению натрий-кальциевых каналов. Сначала ионы натрия, а затем ионы кальция устремляются внутрь клетки миоцита. Этот процесс сопровождается образованием растянутого плато, а функционально фиксируется абсолютная рефрактерность сердечной мышцы.
Эта фаза у рыб значительно продолжительнее — около 0,15 с.
Следующая за этим активизация калиевых каналов и выход ионов калия из клетки обеспечивают быструю реполяризацию мембраны миоцита.
В свою очередь, реполяризация мембраны закрывает калиевые и открывает натриевые каналы. В итоге потенциал клеточной мембраны возвращается к исходному уровню минус 50 мВ.
Миоциты сердца рыбы, способные к генерации потенциала, локализованы в определенных участках сердца, которые совокупно объединены в «проводящую систему сердца». Как и у высших позвоночных, у рыб инициирование сердечной систолы происходит в синатриальном узле.
В отличие от других позвоночных у рыб роль пейсмейкеров выполняют все структуры проводящей системы, которая у костистых включает в себя центр ушкового канала, узел в атриовентрикулярной перегородке, от которого к типичным кардиоцитам желудочка тянутся клетки Пуркинье.
Скорость проведения возбуждения по проводящей системе сердца у рыб ниже, чем у млекопитающих, причем в разных участках сердца она неодинакова.
Максимальная скорость распространения потенциала зарегистрирована в структурах желудочка.
Электрокардиограмма рыб напоминает электрокардиограмму человека в отведениях V3 и V4 (рис. 7.2). Однако техника наложений отведений для рыбы не разработана так подробно, как для наземных позвоночных животных.
Рис. 7.2. Электрокардиограмма рыбы
На электрокардиограмме угря зубцу Р предшествует зубец V. Этиология зубцов такова: зубец Р соответствует возбуждению ушкового канала и сокращению венозного синуса и предсердия; комплекс QRS характеризует возбуждение атриовентрикулярного узла и систолу желудочка; зубец Т возникает в ответ на реполяризацию клеточных мембран сердечного желудочка.
Круг кровообращения рыб
У рыб, как известно, один круг кровообращения. И, тем не менее, кровь по нему циркулирует дольше.
На полный кругооборот крови у рыб уходит около 2 мин (у человека через два круга кровообращения кровь проходит за 20-30 с). Из желудочка через артериальную луковицу или артериальный конус кровь поступает в так называемую брюшную аорту, отходящую от сердца в краниальном направлении к жабрам (рис.7.3).
Брюшная аорта делится на левые и правые (по количеству жаберных дуг) приносящие жаберные артерии. От них к каждому жаберному лепестку отходит лепестковая артерия, а от нее к каждому лепесточку отходят две артериолы, которые формируют капиллярную сеть из тончайших сосудов, стенка которых образована однослойным эпителием с большими Межклеточными пространствами.
Капилляры сливаются в единую выносящую артериолу (по количеству лепесточков). Выносящие артериолы формируют выносящую лепестковую артерию. Лепестковые артерии образуют левую и правую выносящие жаберные артерии, по которым течет артериальная кровь.
Рис. 7.3. Схема кровообращения костистой рыбы
От выносящих жаберных артерий к голове отходят сонные артерии. Далее жаберные артерии сливаются с образованием единого крупного сосуда — спинной аорты, которая тянется по всему телу под позвоночником и обеспечивает артериальное системное кровообращение.
Основными отходящими артериями являются подключичная, брыжеечная, подвздошная, хвостовая и сегментарные. Венозную часть круга начинают капилляры мышц и внутренних органов, которые, объединяясь, формируют парные передние и парные задние кардинальные вены. Кардинальные вены, объединяясь с двумя печеночными венами, образуют кювьеровы протоки, впадающие в венозный синус.
Таким образом, сердце рыб нагнетает и насасывает только венозную кровь.
Однако все органы и ткани получают артериальную кровь, так как перед заполнением микроциркуляторного русла органов кровь проходит через жаберный аппарат, в котором осуществляется обмен газов между венозной кровью и водной средой.
Выделительная система.
Между позвоночником и плавательным пузырем располагаются почки. Образующаяся в них моча по мочеточникам собирается в мочевой пузырь, который открывается наружу отверстием.
Центральная нервная система рыб имеет вид трубки. Передняя часть ее видоизменена в головной мозг, защищенный костями. черепной коробки. В головном мозге позвоночных животных различают пять отделов: передний мозг, промежуточный мозг, средний мозг, мозжечок, продолговатый мозг. У рыб мозг невелик. Наиболее развиты средний мозг и мозжечок.
Ориентацию рыб в воде обеспечивают органы зрения, слуха, обоняния, осязания, вкуса, а также боковая линия, служащая органом чувств, обеспечивающим рыбе ориентацию в воде. Это погруженный в кожу канал, в котором располагаются нервные окончания, воспринимающие давление и направление тока воды. С внешней средой канал сообщается через отверстия в чешуях, покрывающих канал сверху.
Кровеносная система рыб: замкнутая или незамкнутая
Есть ли у рыбы сердце?
В кровеносной системе рыб, по-сравнению с ланцетниками, появляется настоящее сердце. Оно состоит из двух камер, т. е. сердце рыб двухкамерное.
Первая камера — это предсердие, вторая камера — это желудочек сердца. Кровь сначала попадает в предсердие, затем мышечным сокращением проталкивается в желудочек. Далее в результате его сокращения изливается в крупный кровеносный сосуд.
Сердце рыб находится в околосердечной сумке, расположенной за последней парой жаберных дуг в полости тела.
Как и у всех хордовы, кровеносная система рыб замкнутая.
Это значит, что нигде по пути своего следования кровь не покидает сосудов и не изливается в полости тела. Чтобы обеспечить обмен веществ между кровью и клетками всего организма, крупные артерии (сосуды, несущие кровь, насыщенную кислородом) постепенно ветвятся на более мелкие. Самые мелкие сосуды — капилляры. Отдав кислород и забрав углекислый газ, капилляры снова объединяются в более крупные сосуды (но уже венозные).
У рыб только один круг кровообращения.
При двухкамерном сердце по-другому быть и не может. У более высокоорганизованных позвоночных (начиная с земноводных) появляется второй (легочный) круг кровообращения. Но у этих животных и сердце трехкамерное или даже четырехкамерное.
Через сердце протекает венозная кровь, отдавшая кислород клеткам тела.
Далее эту кровь сердце толкает в брюшную аорту, который идет к жабрам и ветвится на приносящие жаберные артерии (но несмотря на название «артерии» они содержат венозную кровь). В жабрах (а конкретно, в жаберных лепестках) из крови в воду выделяется углекислый газ, а из воды в кровь просачивается кислород.
Происходит это в результате разницы в их концентрации (растворенные газы идут туда, где их меньше). Обогатившись кислородом, кровь становится артериальной. Выносящие жаберные артерии (уже с артериальной кровью) впадают в один крупный сосуд — спинную аорту.
Она проходит под позвоночником вдоль тела рыбы и от нее берут начало более мелкие сосуды. От спинной аорты также отходят сонные артерии, идущие к голове и снабжающие кровью в том числе головной мозг.
Перед тем как попасть в сердце венозная кровь проходит через печень, где очищается от вредных веществ.
В кровеносной системе костных и хрящевых рыб есть небольшие различия. В основном это касается сердца. У хрящевых рыб (и некоторых костных) расширенный участок брюшной аорты сокращается наряду с сердцем, а у большинства костных рыб — нет.
Кровь рыб красная, в ней присутствуют эритроциты с гемоглобином, связывающим кислород.
Однако эритроциты рыб имеют овальную форму, а не дисковидную (как, например, у человека). Количество крови, текущей по кровеносной системе, у рыб меньше, чем у наземных позвоночных.
Сердце рыб бьется не часто (около 20-30 ударов в минуту), и количество сокращений зависит от температуры окружающей среды (чем теплее, тем чаще).
Поэтому их кровь течет не так быстро и, следовательно, обмен веществ относительно медленный. Это, например, влияет на то, что рыбы — холоднокровные животные.
У рыб органами кроветворения являются селезенка и соединительная ткань почек.
Несмотря на то, что описанная кровеносная система рыб характерна для подавляющего большинства из них, у двоякодышащих и кистеперых она несколько отличается.
У двоякодышащих в сердце появляется неполная перегородка и появляется подобие легочного (второго) круга кровообращения. Но этот круг проходит не через жабры, а через плавательный пузырь, превращенный в легкое.
Через сердце рыб проходит а) артериальная кровь б) смешанная кровь в) венозная кровь?
Как выглядит сердце рыбы
Сердце рыбы щуки фото.
Есть ли у рыбы сердце, конечно есть.
Фото рыбы щуки с сердцем.
Кровь в сердце рыб проходит также как и у других обеспечивая органы всем для жизни не обходимым.
Сколько сердец у рыбы, речной только одно.
Где у рыбы сердце, в районе гортани и у щуки оно продолжает биться не которое время даже после того как будет извлечено из рыбы.
Какая кровь в сердце рыб, кровь в сердце рыбы щуки такого же красного цвета которая заметно темнеет при чистке.
Фото кровь в сердце рыбы.
Полезные рыбы для сердца практически все речные, только размеры самого сердца слишком малы для употребления в гастрономических целях.
Эта запись была размещена в рыбы автор Vladimir Veselov (постоянная ссылка).
Кровь выполняет многочисленные функции только тогда, когда движется по сосудам. Обмен веществ между кровью и другими тканями организма происходит в капиллярной сети. Отличаясь большой протяженностью и разветвленностью, она оказывает большое сопротивление току крови. Давление, необходимое для преодоления сопротивления сосудов, создается в основном сердцем, Строение сердца рыб проще, чем высших позвоночных. Производительность сердца у рыб как нагнетательного насоса значительно ниже, чем у наземных животных.
Тем не менее оно справляется со своими задачами. Водная среда создает благоприятные условия для работы сердца. Если у наземных животных значительная часть работы сердца затрачивается на преодоление сил гравитации, вертикальные перемещения крови, то у рыб плотная водная среда существенно нивелирует гравитационные влияния.
Вытянутое в горизонтальном направлении тело, небольшой объем крови, наличие только одного крута кровообращения дополнительно облегчают функции сердца у рыб.
Строение сердца рыб
Сердце у рыб небольшое, составляющее примерно 0,1% массы тела. Из этого правила, конечно, есть исключения. Например, у летучих рыб масса сердца достигает 2,5 % массы тела.
Для всех рыб характерно двухкамерное сердце. Вместе с тем существуют видовые различия в строении этого органа.
В обобщенном виде можно представить две схемы строения сердца в классе рыб. И в первом, и во втором случае выделяют 4 полости: венозный синус, предсердие, желудочек и образование, отдаленно напоминающее дугу аорты у теплокровных, — артериальную луковицу у костистых и артериальный конус у пластинчатожаберных (рис, 7.1). Принципиальное различие этих схем заключено в морфофункциональных особенностях желудочков и артериальных образований.
У костистых артериальная луковица представлена фиброзной тканью с губчатым строением внутреннего слоя, но без клапанов.
У пластинчатожаберных артериальный конус помимо фиброзной ткани содержит и типичную сердечную мышечную ткань, поэтому обладает сократимостью.
Конус имеет систему клапанов, облегчающих одностороннее продвижение крови через сердце.
 |
Рис. 7.1. Схема строения сердца рыб
В желудочке сердца рыб обнаружены различия в структуре миокарда.
Принято считать, что миокард рыб специфичен и представлен однородной сердечной тканью, равномерно пронизанной трабекулами и капиллярами. Диаметр мышечных волокон у рыб меньше, чем у теплокровных, и составляет 6-7 мкм, что вдвое меньше по сравнению, например, с миокардом собаки. Такой миокард называют губчатым.
Сообщения о васкуляризации миокарда рыб довольно запутанны. Миокард снабжается венозной кровью из трабекулярных полостей, которые, в свою очередь, заполняются кровью из желудочка через сосуды Тибезия (Thebesian vessels). В классическом понимании у рыб нет коронарного кровообращения. По крайней мере, медики-кардиологи придерживаются такой точки зрения. Однако в литературе по ихтиологии термин «коронарное кровообращение рыб» встречается часто.
В последние годы исследователи обнаружили много вариаций васкуляризации миокарда. Например, С. Agnisola et. al (1994) сообщает о наличии двуслойного миокарда у форели и электрического ската. Со стороны эндокарда лежит губчатый слой, а над ним слой миокардиальных волокон с компактным упорядоченным расположением.
Исследования показали, что губчатый слой миокарда обеспечивается венозной кровью из трабекулярных лакун, а компактный слой получает артериальную кровь по гипобронхиальным артериям второй пары жаберных дут.
У elasmobranchs коронарное кровообращение отличается тем, что артериальная кровь из гипобронхиальных артерий доходит до губчатого слоя по хорошо развитой системе капилляров и попадает в полость желудочка по сосудам Тибезия.
Еще одно существенное различие костистых и пластинчатожаберных заключается в морфологии перикарда.
У костистых перикард напоминает таковой наземных животных. Он представлен тонкой оболочкой.
У пластинчатожаберных перикард образован хрящевой тканью поэтому он представляет собой как бы жесткую, но упругую капсулу.
В последнем случае в период диастолы в перикардиальном пространстве создается некоторое разрежение, что облегчает кровенаполнение венозного синуса и предсердия без дополнительных затрат энергии.
Электрические свойства сердца рыб
Строение миоцитов сердечной мышцы рыб сходно с таковым высших позвоночных.
Поэтому и электрические свойства сердца похожи. Потенциал покоя миоцитов у костистых и пластинчатожаберных составляет 70 мВ, у миксин — 50 мВ. На пике потенциала действия регистрируется изменение знака и величины потенциала с минус 50 мВ до плюс 15 мВ. Деполяризация мембраны миоцита приводит к возбуждению натрий-кальциевых каналов. Сначала ионы натрия, а затем ионы кальция устремляются внутрь клетки миоцита. Этот процесс сопровождается образованием растянутого плато, а функционально фиксируется абсолютная рефрактерность сердечной мышцы.
Эта фаза у рыб значительно продолжительнее — около 0,15 с.
Следующая за этим активизация калиевых каналов и выход ионов калия из клетки обеспечивают быструю реполяризацию мембраны миоцита.
В свою очередь, реполяризация мембраны закрывает калиевые и открывает натриевые каналы. В итоге потенциал клеточной мембраны возвращается к исходному уровню минус 50 мВ.
Миоциты сердца рыбы, способные к генерации потенциала, локализованы в определенных участках сердца, которые совокупно объединены в «проводящую систему сердца». Как и у высших позвоночных, у рыб инициирование сердечной систолы происходит в синатриальном узле.
В отличие от других позвоночных у рыб роль пейсмейкеров выполняют все структуры проводящей системы, которая у костистых включает в себя центр ушкового канала, узел в атриовентрикулярной перегородке, от которого к типичным кардиоцитам желудочка тянутся клетки Пуркинье.
Скорость проведения возбуждения по проводящей системе сердца у рыб ниже, чем у млекопитающих, причем в разных участках сердца она неодинакова.
Максимальная скорость распространения потенциала зарегистрирована в структурах желудочка.
Электрокардиограмма рыб напоминает электрокардиограмму человека в отведениях V3 и V4 (рис. 7.2). Однако техника наложений отведений для рыбы не разработана так подробно, как для наземных позвоночных животных.
 |
Рис. 7.2. Электрокардиограмма рыбы
На электрокардиограмме угря зубцу Р предшествует зубец V. Этиология зубцов такова: зубец Р соответствует возбуждению ушкового канала и сокращению венозного синуса и предсердия; комплекс QRS характеризует возбуждение атриовентрикулярного узла и систолу желудочка; зубец Т возникает в ответ на реполяризацию клеточных мембран сердечного желудочка.
Работа сердца рыб
Сердце рыб работает ритмично.
Частота сердечных сокращений у рыб зависит от многих факторов.
Частота сердечных сокращений (ударов в минуту) у карпа при 20 °С
Молодь массой 0,02 г 80
Сеголетки массой 25 г 40
Двухлетки массой 500 г 30
В опытах in vitro (изолированное перфузированное сердце) частота сердечных сокращений у радужной форели и электрического ската составила 20-40 ударов в минуту.
Из множества факторов наиболее выраженное влияние на частоту сердечных сокращений оказывает температура среды обитания.
Методом телеметрии на морском окуне и камбале была выявлена следующая зависимость (табл. 7.1).
| Температура, °С | Частота сердечных сокращений, ударов в минуту | Температура, °С | Частота сердечных сокращений, ударов в минуту |
| 24 | 11,5 | 31 | |
| 26 | 43 | ||
| 29 |
Установлена видовая чувствительность рыб к перепадам температуры.
Так, у камбалы при повышении температуры воды с g до 12 аС частота сердечных сокращений возрастает в 2 раза (с 24 до 50 ударов в минуту), у окуня — только с 30 до 36 ударов в минуту.
Регуляция сердечных сокращений осуществляется при помощи центральной нервной системы, а также внутрисердечных механизмов.
Интересно, что один и тот же гуморальный агент при различной температуре окружающей среды может оказывать прямо противоположное воздействие на сердце рыб.
Так, на изолированном сердце форели при низких температурах (6аС) эпинефрин вызывает положительный хронотропный эффект, а на фоне повышенных температур (15аС) перфузирующей жидкости — отрицательный хронотропный эффект.
Сердечный выброс крови у рыб оценивается в 15-30 мл/кг в минуту. Линейная скорость крови в брюшной аорте составляет 8- 20 см/с.
In vitro на форели установлена зависимость сердечного выброса от давления перфузирующей жидкости и содержания в ней кислорода. Однако в тех же условиях у электрического ската минутный объем не изменялся. В состав перфузата исследователи включают более десятка компонентов.
Состав перфузата для сердца форели (г/л)
Фторид кальция 0,23
Сульфат магния (кристаллический) 0,23
Фосфат натрия однозамещенный (кристаллический) 0,016
Фосфат натрия двузамещенный (кристаллический) 0,41
Поливинил пиррол идол (PVP) коллоидный 10,0
Раствор насыщается газовой смесью из 99,5 % кислорода, 0,5 % углекислого газа (диоксида углерода) или смесью воздуха (99 5%) с углекислым газом (0,5 %).
2. рН перфузата доводят до 7,9 при температуре 10 аС, используя бикарбонат натрия.
Состав перфузата для сердца электрического ската (г/л)
Хлорид натрия 16,36
Сульфат натрия 0,071
Фосфат натрия однозамещенный (кристаллический) 0,14
Бикарбонат натрия 0,64
Перфузат насыщают той же газовой смесью. 2.рН 7,6.
В таких растворах изолированное сердце рыб сохраняет физиологические свойства и функционирует очень долго. При выполнении простых манипуляций с сердцем допускается использование изотонического раствора хлорида натрия. Однако не стоит при этом рассчитывать на продолжительную работу сердечной мышцы.
Круг кровообращения рыб
У рыб, как известно, один круг кровообращения. И, тем не менее, кровь по нему циркулирует дольше.
На полный кругооборот крови у рыб уходит около 2 мин (у человека через два круга кровообращения кровь проходит за 20-30 с). Из желудочка через артериальную луковицу или артериальный конус кровь поступает в так называемую брюшную аорту, отходящую от сердца в краниальном направлении к жабрам (рис.7.3).
Брюшная аорта делится на левые и правые (по количеству жаберных дуг) приносящие жаберные артерии. От них к каждому жаберному лепестку отходит лепестковая артерия, а от нее к каждому лепесточку отходят две артериолы, которые формируют капиллярную сеть из тончайших сосудов, стенка которых образована однослойным эпителием с большими Межклеточными пространствами.
Капилляры сливаются в единую выносящую артериолу (по количеству лепесточков). Выносящие артериолы формируют выносящую лепестковую артерию. Лепестковые артерии образуют левую и правую выносящие жаберные артерии, по которым течет артериальная кровь.
Рис. 7.3. Схема кровообращения костистой рыбы
От выносящих жаберных артерий к голове отходят сонные артерии. Далее жаберные артерии сливаются с образованием единого крупного сосуда — спинной аорты, которая тянется по всему телу под позвоночником и обеспечивает артериальное системное кровообращение.
Основными отходящими артериями являются подключичная, брыжеечная, подвздошная, хвостовая и сегментарные. Венозную часть круга начинают капилляры мышц и внутренних органов, которые, объединяясь, формируют парные передние и парные задние кардинальные вены. Кардинальные вены, объединяясь с двумя печеночными венами, образуют кювьеровы протоки, впадающие в венозный синус.
Таким образом, сердце рыб нагнетает и насасывает только венозную кровь.
Однако все органы и ткани получают артериальную кровь, так как перед заполнением микроциркуляторного русла органов кровь проходит через жаберный аппарат, в котором осуществляется обмен газов между венозной кровью и водной средой.
Движение крови и кровяное давление у рыб
Кровь движется по сосудам вследствие разницы ее давления в начале круга кровообращения и в его конце.
При измерении кровяного давления без анестезии в вентральном положении (вызывает брадикардию) у лосося в брюшной аорте оно составило 82/50 мм рт. ст., а в дорзальной 44/37 мм рт. ст. Исследование анестезированных рыб нескольких видов показало, что анестезия существенно снижает систолическое давление — до 30-70 мм рт. ст.
Пульсовое давление при этом по видам рыб колебалось от 10 до 30 мм рт. ст. Гипоксия приводила к повышению пульсового давления до 40 мм рт. ст.
В конце круга кровообращения давление крови на стенки сосудов (в кювьеровых протоках) не превышало 10 мм рт. ст.
Наибольшее сопротивление току крови оказывает жаберная система с ее длинными и сильно разветвленными капиллярами.
У карпа и форели разница систолического давления в брюшной и дорзальной аортах, т. е. при входе и на выходе из жаберного аппарата, составляет 40-50 %. При гипоксии жабры оказывают еще большее сопротивление току крови.
Помимо сердца продвижению крови по сосудам способствуют и другие механизмы.
Так, дорзальная аорта, имеющая форму прямой трубы со сравнительно жесткими (по сравнению с брюшной аортой) стенками, оказывает незначительное сопротивление току крови. Сегментарная, каудальная и другие артерии имеют систему кармашковых клапанов, аналогичную тем, которые есть у крупных венозных сосудов.
Эта система клапанов препятствует обратному току крови. Для венозного тока крови большое значение имеют также сокращения прилегающих к венам мыши, которые проталкивают кровь в кардиальном направлении. Венозный возврат и сердечный выброс оптимизируются мобилизацией депонированной крови. Экспериментально доказано, что у форели мышечная нагрузка приводит к уменьшению объема селезенки и печени.



